免疫应答作为一种生理功能,无论是对自身成分的耐受现象,还是对“非已”抗原的排斥都是机体的免疫调节机制的控制下进行的。免疫调节机制是维持机体内环境稳定的关键,如果免疫调节功能异常,对自身成分产生强烈的免疫攻击,造成细胞破坏,功能丧失,就会发生自身免疫病。如果对外界病原微生物感染不能产生适度的反应(反应过低可造严重感染,反应过强则发生过敏反应),也可造成对机体的有害作用。因此,免疫调节机制不仅决定了免疫应答的发生,而且也决定了反应的强弱。这一调节作用是精细的、复杂的。调节功能是作用于免疫应答过程中的多个环节。
第一节 免疫系统的调节
一、抗原的调节作用
免疫应答的发生是由抗原刺激免疫系统的细胞开始的。抗原的存在是应答发生的前提,但是抗原在体内可被分解代谢,而浓度逐渐降低,抗体产生也不断下降。已有实验证明,在免疫应答过程中,由于注射大量抗体,清除了抗原,导致抗体分泌细胞数量减少,抗体合成也明显下降。
二、抗体的调节作用
(一)抗体的反馈调节作用
抗体对免疫应答也有反馈调节作用。抗体是免疫应答的产物,抗体产生之后又可抑制其后的抗体产生。将抗体注入非免疫的机体,可阻止其后注入抗原引起的免疫应答,这一现象在临床上应用成功地预防了新生儿溶血性疾病的发生。该疾病是因母子间RH血型不符引起的。应用抗Rh因子抗体给分娩Rh+胎儿的Rh-产妇注射,由于分娩过程中进入母体Rh抗原被注入的Rh抗体所清除,因而抑制了Rh母亲进一步产生抗体,也就防止了因Rh抗体(IgG)通过胎盘使下次妊娠的Rh+胎儿产生溶血症。
免疫应答过程中用血浆交换法去除循环血中的抗体,使血液中的抗体浓度不断降低,可促使抗体产生增加。注射IgG抗体可明显促使抗体形成细胞数量下降,可反馈控制抗体过度合成,这可能是由于抗体结合抗原后降低抗原的刺激作用。
(二)抗原抗体复合物的调节作用
抗原抗体复合物也有调节免疫应答的作用。免疫复合物抗原可与B细胞表面的抗原受体结合,复合物中的抗体可与B细胞表面的Fc受体结合,当B细胞表面的抗原受体的Fc受体因抗原抗体复合物的作用而发生交联时,就可产生抑制信号,可抑制B细胞分化为抗体形成细胞。但当抗原量多,抗体量少时形成的复合物可与抗原呈递细胞(APC)表面的Fc受体结合,则可增强抗原呈递细胞的功能,进而增强B细胞产生抗体的反应。所以免疫复合物的调节作用在反应初期由于抗原量大,多表现为增强反应,而到后期由于抗体量增多可中和抗原而起抑制作用。此外,抗体类别不同,作用也不同,在反应初期,当IgM产生多时,形成的复合物有增强作用,而后期IgG多时,则起抑制作用(图14-2)
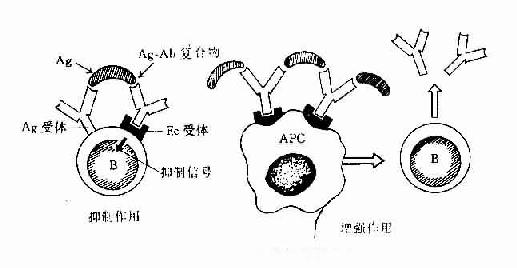
图14-1 免疫复合物的免疫调节作用
三、免疫细胞的调节作用
T细胞和单核-巨噬细胞既是免疫应答的效应细胞,也参予免疫应答的调节。
(一)TH细胞的调节作用
抗原刺激在机体内发生的体液免疫或细胞免疫都是由抗原呈递细胞和T辅助细胞(TH)的相互作用开始的。TH细胞因分泌细胞因子种类不同而分成TH1和TH2两类,诱发体液免疫作用的是TH2,诱发细胞免疫起辅助作用的是TH1。
TH1和TH2还可通过各自分泌的细胞因子相互制约。TH1细胞主要产生IL-2和IFN-r,后者可抑制TH2增殖和功能,而TH2产生IL-4、IL-5和IL-10,特别是IL-10是个重要的免疫抑制因子,它可抑制TH1分泌IL-2和IFN-r,抑制MHCⅡ类分子的表达。并可抑制Mф产生IL-1、IL-6等。
(二)TS细胞的调节作用
在免疫应答过程中,经辅助性诱导T细胞(CD4+T)的作用可活化抑制性T细胞(TS),使之分化成为效应TS细胞。它可分泌抗原特异及非特异抑制因子,可抑制杀伤性T细胞(TC)、辅助性T细胞(TH)及B细胞的功能,发挥负反馈调节作用。如此,当外来抗原侵入机体后经APC活化TH细胞以后启动正免疫应答,产生效应分子和(或)效应细胞以清除外来抗原。与此同时在免疫应答后期可启动TS细胞使之活化及分化发育为效应TS细胞,以抑制免疫应答,籍以维持机体的免疫稳定平衡。
(三)TC(CTL)细胞的调节作用
T细胞抑制免疫应答也可通TC的作用,因为TC可针对T或B细胞表面TCR或BCR可变区的独特型决定基而起特异杀伤作用。由于TC细胞杀伤T或B细胞而引起免疫抑制作用。
四、独特型网络调节
(一)免疫网络学说的提出
这一学说是Jerne(1972)根据现代免疫学对抗体分子独特型的认识而提出的。这一学说认为在抗原刺激发生之前,机体处于一种相对的免疫稳定状态,当抗原进入机体后打破了这种平衡,导致了特异抗体分子的产生,当达到一定量时将引起抗Ig分子独特型的免疫应答,即抗抗体的产生。因此抗体分子在识别抗原的同时,也能被其它抗体分子所识别。这一点无论对血流中的抗体分子或是存在于淋巴细胞表面作为抗原受体的Ig分子都是一样的。在同一动物体内一组抗体分子亦可被另一组淋巴细胞表面抗独特型抗体分子所识别。而一组淋巴细胞表面抗原受体分子亦可被另一组淋巴细胞表面抗独特型抗体分子所识别。这样在体内应形成了淋巴细胞与抗体分子所组成的网络结构。网络学说认为,这种抗抗体的产生在免疫应答的调节中起着重要作用。使受抗原刺激增殖的克隆受到抑制,而不至于无休止地进行增殖。籍以维持免疫应答的稳定平衡。
(二)独特型网络的细胞
独特型决定簇存在于Ig的V区,也可存在于各类T细胞及B细胞的抗原识别受体的V区。因此在体内形成由独特型(idiotype,Id)和抗独特型(antiId)组成的免疫网络。
就淋巴细胞来说,构成这种网络结构的淋巴细胞有四种类型。当抗原进入机体后可与相应的抗原反应细胞相结合,进行增殖、分化并产生抗体分子。这种抗原反应细胞可与另外三种淋巴细胞构成网络。一组是独特型反应细胞,即抗独特型淋巴细胞组,能识别抗原受体的独特型,具有抑制抗原反应淋巴细胞的作用,另一组能增强抗原反应淋巴细胞的作用,它的受体带有与抗原构型相同的独特型,因此也能被抗原反应细胞所识别,Jerne称此组淋巴细胞为内影像组。内影像概念是免疫网络理论的重要组成部分。第三组淋巴细胞为非特异平行组,其抗原识别受体与抗原反应细胞不同,但独特型却与之相同,本组细胞可促进独特型细胞的活性,可加强对网络的抑制作用。同样这三组淋巴细胞也各自通过其独特型的联系和其它淋巴细胞也形成网络,如此不断扩展。所以机体对某一特定抗原应答不只表现为抗原反应细胞的应答,而是通过独特型联结起来的一个庞大的免疫网络整体反应,它们通过连续不断的识别过程,产生促进或抑制作用,以维持机体免疫应答的相对稳定状态(图14-2)。
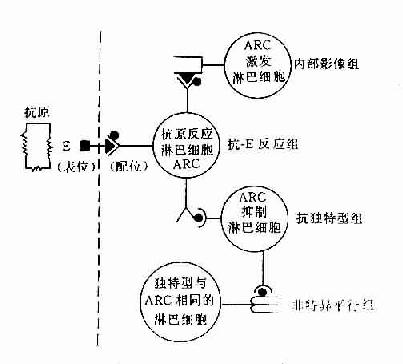
图14-2 免疫网络学说
(三)独特型网络理论的应用意义
独特型理论为人工调控免疫应答提供新的思路,特别是处于超敏状态下如过敏症、自身免疫病和器官移植时,已有利用抗Id抗体的抑制作用进行实验治疗成功的报导。例如用B大鼠的移植抗原注射A大鼠,自A品系大鼠获得抗体,用此抗体对A大鼠进行免疫产生抗Id抗体就可抑制由T细胞介导的对B移植抗原的排斥反应。这可能因为抗-Id反应灭活了引起排斥反应的淋巴细胞,也就是抗-Id封闭了B细胞受体上的Id。另外一种完全不同的方法是应用抗原内影像的抗-Id刺激抗原特异T抑制细胞活化,能阻断对同一抗原上对其他抗原决定簇起反应的B细胞活化,这也就是抗原本身的桥梁作用。
某些情况下也可应用抗原内影像即抗-Id代替抗原刺激产生抗体(图14-3)。这种情况用于抗原数量少,难以获得时,如某些寄生虫抗原、某些癌相关的胚胎抗原、用化学合成方法得到的抗原或用基因克隆法得到的重组抗原难以折叠成天然分子构型的蛋白质抗原,选择有抗原表位(epiope)构型的抗-Id代替抗原进行免疫以克服抗原不足的困难。
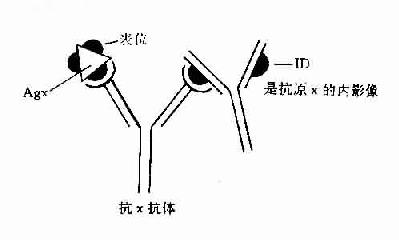
图14-3 内影像图解
第二节 遗传对免疫应答的调节
一、高应答品系与低应品系的产生
小鼠可选择性的培育出产生高或低抗体应答的品系,经过几代培养,一些品系可以稳定的对多种抗原产生高滴度抗体,而另一些品系则可稳定的产生低水平的抗体(图14-4)。
经研究证明上述高或低应答小鼠,至少有10个基因座和这种反应高低有关,其中主要是影响Mф特性基因,观察Mф从血液中清除碳颗粒或吞噬SRBC的能力来比较两品系小鼠Mф功能的不同,发现高应答品系小鼠的Mф功能活性好,有较多的细胞表面经处理过的抗原,而低应答品系小鼠的Mф相反。
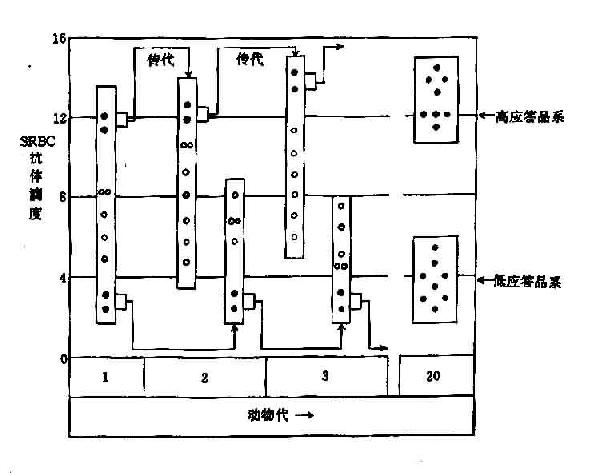
图14-4 高或低应答品系小鼠培养过程
用SRBC免疫一组小鼠,SRBC是多决定簇抗原。图中每支小鼠的抗体滴度由圆圈表示,
选出Ab滴度最高的雌雄交配繁育后代。用抗原再免疫,同样再选出滴度最高的一对繁育后代。经过20代后,所有后代均是对SRBC产生高滴度抗体,同时对其他多种抗原也是高抗体应答,同样方法也可选育出低应答品系小鼠
二、MHC(Ir基因)对免疫应答的影响
已发现对多种简单的TD抗原的应答能力是由MHC的基因决定的。例如H-2b单体型(haplotype)小鼠对合成多肽抗原(T,G)-A-L反应很好,而H-2k小鼠则反应很差,产生抗体很少。H-2b单体型小鼠(具有特殊H-2基因)对(T,G)-A-L抗原是高应答小鼠,因为它们具有适宜Ir基因。对另一种合成抗原(G,G)-A-L,是用组氨酸(H)代替了酪氨酸(T),反应情况则完全相反,原来对(T,G)-A-L反应差的对(H,G)-A-L反应很好,说明各种不同系小鼠应答能力高低不但与Ir基因结构有关,也与抗原构造有关(表14-1)。
表14-1 H-2单倍型对合成多肽抗原的应答
抗原 |
H-2单倍型 |
B |
K |
D |
A |
S |
(T,G)-A-L |
高 |
低 |
中 |
低 |
低 |
(H,G)-A-L |
低 |
高 |
中 |
高 |
低 |
这种H-2单倍型与应答能力高低之间的关系仅仅是用结构明确、构造简单的抗原进行研究的结果。因为对这些简单抗原决定簇的应答是由Ir基因控制的。
三、Ir基因控制T-B细胞间的协作
用几种基因重组小鼠品系研究对简单合成抗原的应答,见有14-2。
表14-2 不同品系小鼠Ir基因与免疫应答的关系
小鼠品系 |
H-2区 |
对(H,G)-A-L |
K |
IA |
I-E |
S |
D |
应答 |
A |
k |
k |
k |
b |
b |
高 |
A.TL |
s |
k |
k |
k |
b |
高 |
B.10.A(4R) |
k |
k |
b |
b |
b |
高 |
B.10 |
B |
b |
b |
b |
b |
低 |
A.SW |
s |
s |
s |
s |
s |
低 |
表中三个高反应品系小鼠具有相同的I-AK和H-2Db,而B.10小鼠只有H-2Db,它是低反应的。所以高反应必然与I-Ak有关系。由於小鼠的I-A亚区基因的点突变,导致基因编码的MHCⅡ类分子改变(也就是影响Ⅱ类分子多形特异部位)从而改变小鼠从高应答变成低应答状态。I-A亚区基因点突变也大大降低了T细胞增殖能力。抗原特异T细胞增殖能力与宿主应答状态有密切关系,即高应答动物的T细胞增殖能力也强,反之则弱。
第三节 神经内分泌免疫网络调节
一、神经内泌对免疫系统的调节
已证明免疫细胞上有接受神经递质和激素刺激的受体,可以说几乎所有的免疫细胞上都有不同的神经递质及内分泌激素的受体。这些内分泌激素和神经递质都具有免疫调节功能,如肾上肾上腺皮质激素,它是最早发现的具有调节免疫功能的激素,它几乎对所有的通过免疫细胞都的抑制作用,包括淋巴细胞、巨噬细胞、中性粒细胞和肥大细胞。刺激下丘脑可通过促肾上腺皮质激素释放因子(CRF)引起垂体释放ACTH,通过血液循ACTH可促进肾上腺皮质释放糖皮质激素,因而形成下丘脑-垂体-肾上腺轴。各种应激刺激可以使血中肾上腺皮质激素含量增高,所以由应激引起免疫抑制与这类激素的作用有密切关系。除此之外还存非垂体-肾上腺轴的免疫调节作用,这包括生长激素、生乳素和阿片肽等的作用。其中已证明生长激素和生乳素对多种免疫细胞有促分化和增强功能的作用,而阿片肽对免疫细胞影响增强和抑制的报导均有,作用复杂,有待深入研究。
二、免疫系统对神经内泌系统的调节作用
免疫系统可以通过多种途径影响神经内分泌功能。免疫细胞本身可以产生和释放内分泌激素,也可以通过它们所产生的细胞因子作用于神经内分泌及全身各器官系统。
(一)免疫细胞产生的内分泌激素
1980年Blalocd等证明人白细胞干扰素中有ACTH和r-内啡肽的活性片段。他们用胃蛋白酶消化人白细胞干扰素,可得到具有ACTH活性的小分子片段。经受体分析也证明,人IFN-α能与与3H双氢吗啡竞争受体,且比吗啡强300倍以上。他们用抗ACTH和r-内啡肽的荧光抗体染色,发现人外周血淋巴几乎100%呈现阳性。这些结果说明免疫细胞可以产生某些内分泌激素。此外,Zurawski在小鼠T辅助细胞株中找到啡肽前体的mRNA(preproenkephalin mRNA)在cDNA文中可达0.4%。以ConA)刺激T辅助细胞后清液中可以测到脑啡肽样免疫活性物质。这一发现从基因表达证明小鼠TH细胞可以合成和释放脑啡肽。目前已发现免疫细胞合成的神经递质和激素达10余种(表14-3)。
表14-3 免疫细胞产生的神经内分泌肽
名称 |
产生细胞 |
名称 |
产生细胞 |
ACTH |
淋巴细胞和巨噬细胞性 |
生长抑制素 |
单核细胞、肥大细胞、多形核白细胞 |
内啡肽 |
淋巴细胞、巨噬细胞 |
脑啡肽 |
辅助性T细胞 |
生长激素 |
淋巴细胞 |
精氨酸升压素(AVP) |
胸腺上皮细胞 |
生乳素 |
淋巴细胞 |
催产素 |
胸腺上皮细胞 |
绒毛膜促性腺素 |
T细胞 |
神经垂体激素运载蛋白 |
|
血管活性肠肽(VIP) |
单核细胞、肥大细胞、多形核白细胞 |
TSH |
胸腺上皮细胞、T细胞 |
(二)免疫细胞产生的细胞因子对神经内分泌系统的作用
免疫细胞产生的淋巴因子和单核因子除对自身活动进行调节外,还可作用到神经内分泌系统,从而影响全身系统的功能活动,其中报导较多的有IL-1、IL-2及干扰素等。
IL-1可刺激下丘脑使体温升高,故又称为内源性致热原。神经系统内星形胶质细胞和小胶质细胞也可产生IL-1,在神经细胞发育和修复中发挥作用。IL-1还可引起慢波睡眠,在外周,它可抑制食欲。机体在对抗原刺激发生反应高峰阶段,血液中糖皮质激素增加而出现抑制免疫的作用。这可能由于IL-1作用于垂体,通过ACTH促使肾上腺皮质激水平升高引起的。
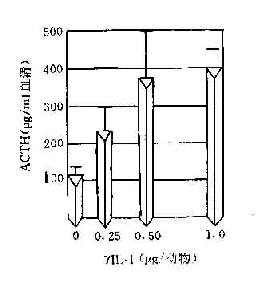
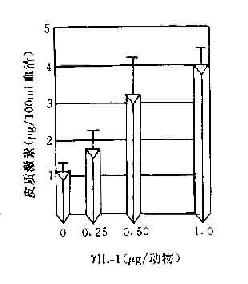
图14-5 由于IL-1和淋巴因子产生使糖皮质激素含量增加而产生免疫抑制
