游离于血液中的单核细胞(monocyte)及存在于体腔和各种组织中的巨噬细胞(macrophage,MФ)均来源于骨髓干细胞,它们具有很强的吞噬能力,且细胞核不分叶,故命名为单核吞噬细胞系统(mononuclear phagocyte system,MPS)。单核/巨噬细胞是一类主要的抗原呈递细胞,在特异性免疫应答的诱导与调节中起着关键的作用。
第一节 单核吞噬细胞系统
一、单核吞噬细胞系统细胞的来源与分化发育
MPS细胞起源于骨髓,其分化与更新受细胞因子复杂网络的调控。在某些细胞因子,如多集落刺激因子(multi-colony stimulating factor,multi-CSF)、巨噬细胞集落刺激因子(macrophage-CSF,GM-CSF)等的刺激下,骨髓中的髓样干细胞经原单核细胞(monoblast)、前单核细胞(pre-monocyte)分化发育为单核细胞并进入血流。外周血单核细胞占白细胞总数1%~3%,它在血流中仅存留几小时至数十小时,然后粘附到毛细血管内皮,穿过内皮细胞接合处,移行至全身各组织并发育成熟为巨噬细胞。组织损伤和炎症可加速单核细胞向组织移行。巨噬细胞在组织中寿命可达数月至数年。在不同组织中存留的巨噬细胞由于局部微环境的差异,其形态及生物学特征均有所不同,名称也各异(表9-1)。一般认为除少数单核细胞或低分化的巨噬细胞外,成熟的巨噬细胞很少有或没有增殖能力,并不断被骨髓前体细胞分化的细胞所补充。另外,单核吞噬细胞系统细胞的分化发育还可受各种细胞因子如IL-2、IL-4以及干扰素等影响。
表9-1 单核吞噬细胞系统细胞的发生和分布
骨髓 |
血液 组织 |
多能干细胞 ↓ 髓样干细胞 ↓ 单核干细胞 ↓ 前单核细胞 ↓ 单核细胞 |
→单核细胞→巨噬细胞→结缔组织:组织细胞 肺:肺泡巨噬细胞 肝:枯否(Kupffer)细胞 脾与淋巴结:游走与固定巨噬细胞 浆膜腔:胸、腹腔巨噬细胞 神经组织:小胶质细胞 骨:破骨细胞 关节:滑膜A型细胞 |
二、单核吞噬细胞系统细胞的解剖学特征
(一)形态结构
单核细胞一般为圆形,直径约10~20μm;巨噬细胞大小不等,直径约10~30μm或更大,常有伪足,呈多形性。单核/巨噬细胞有圆形或椭圆形的核,胞浆中富含溶酶体及其他各种细胞器。
(二)单核吞噬细胞系统细胞的表面分子
1.表面受体 MPS细胞表面有多达80种以上受体分子,它们与相应的配体结合,分别表现感应与效应功能。包括捕获病原异物,加强调理、趋化、免疫粘连、吞噬、介导细胞毒作用等。例如,免疫球蛋白Fc受体(FcγRⅠ即CD64、FcγRⅡ即CD32、FcγRⅢ即CD16)补体受体(CD1即CD35、CD3即CD11b/18或Mac-1)可以分别与IgG的Fc段及补体C3b片段结合,从而促进单核/巨噬细胞的活化和调理吞噬功能。此外,单核/巨噬细胞还表达各种细胞因子、激素、神经肽、多糖、糖蛋白、脂蛋白及脂多糖的受体,从而可感应多种调控其功能的刺激信号。
2.表面抗原 MPS细胞表面具有多种抗原分子,它们对MPS细胞的鉴定与功能有重要意义。例如,MPS细胞表达MHC抗原,尤其MHC-Ⅱ类抗原是巨噬细胞发挥抗原呈递作用的关键性效应分子;单核/巨噬细胞还表达多种粘附分子(abhesionmolecule),如选择素L(L-selectin)、细胞间粘附分子(intercellular adhesion molecule,ICAM)和血管细胞粘附分子(vascylar cell adhesion molecule,VCAM)等,它们介导MPS细胞与其他细胞或外基质间的粘附作用,从而参与炎症与免疫应答过程。近年来应用单克隆抗体鉴定出许多单核吞噬细胞的表面分化抗原,如OKM-1、Mac-120、MO1~4等,但这些抗原也可能表达在其他起源于髓样干细胞的细胞(如中性粒细胞)表面。另外,成熟的单核细胞可表达高密度的CD14,这是一种相对特异的单核细胞表面标志。
三、单核吞噬细胞系统细胞的生理特点
(一)一般性质
MPS细胞又称大吞噬细胞,它具有吞噬细胞的一般特征,如何通过吞饮摄入液体异物,也可通过吞噬摄取颗粒性异物,还可识别某些化学刺激物的浓度,表现出定向运动的能力,即具有趋化性。MPS细胞在吞噬异物后,细胞内会发生一系列代谢改变,如糖代射增强,能量产生增加,活性氧生成增多等。MPS细胞胞浆中还含有非特异性酯酶、碱性磷酸二酯和过氧化物酶等。在细胞分化和激活过程中,这些酶的量及细胞内的定位可发生改变。此外,由于MPS细胞表达丰富的粘附分子,对玻璃与塑料制品具有强的粘附性,故又被称为粘附细胞(adherentcell),借助这个特性可将MPS细胞与淋巴细胞分离。
(二)单核吞噬细胞系统细胞的激活
MPS细胞在环境因素刺激下,可发生形态、膜分子表达以及细胞代谢与功能的短暂、可逆性变化,这一过程称为MPS细胞的激活,也是它有别于其他吞噬细胞(如中性粒细胞)的一个重要特征。与分化过程不同,活化是在病理条件下表现出的可逆性功能状态。单核吞噬细胞的激活是一个复杂的多步骤过程,在不同的活化阶段,涉及不同刺激因子的作用,细胞形态及功能也发生相应的改变。以巨噬细胞(Mф)为例,体内的Mф一般处于静止状态,病原体等异物通过直接接触激发Mф内的生理生化反应。环核苷酸第二信使cAmp/cGMP水平升高,使静止态Mф转变为应答性Mф。后者在IFN-γ等刺激因子启动下转变为致敏的Mф,然后在脂多糖或某些细胞因子作用下转变为活化的Mф。在上述变化过程中,Mф表现出形态改变(浆膜呈不规则波浪形,细胞器增加,膜分子表达改变),代谢增强(胞内蛋白质合成与ATP生成增加,磷酸戊糖代谢增强)以及功能增强(吞噬率及吞噬速度增高,杀菌及杀瘤能力增强,分泌活性及抗原呈递能力增强)等(表9-2)。
表9-2 巨噬细胞的激活
|
静止Mф→ |
病原体 应答性Mф→ |
IFN-γ 致敏的Mф→ |
LPS等 活化Mф |
吞噬功能 |
+ |
++ |
+++ |
++++ |
趋化 |
+ |
++ |
+++ |
++++ |
杀瘤活性 |
- |
- |
- |
+ |
抗原呈递 |
- |
- |
+ |
+ |
TNF分泌 |
- |
- |
- |
+ |
活性氧产生 |
- |
+ |
++ |
++ |
MHC-Ⅱ分子 |
- |
- |
+ |
+ |
转铁蛋白受体 |
- |
+ |
- |
- |
IFN-γ:γ干扰素
LPS:脂多糖
TFN:肿瘤坏死因子
一般认为,只有激活的单核吞噬细胞才是具有活跃生物学作用的效应细胞。在病理情况下,MPS细胞的异常激活也参与某些疾病的发生与发展。
(三)单核吞噬细胞系统细胞的分泌活性
MPS细胞是一类重要的分泌细胞。在许多组织和器官中,MPS细胞是分泌性蛋白的主要来源,其分泌物种类之多在体内仅肝细胞才能与之相比。一般情况下,活化的MPS细胞才有活跃的分泌能力。现已发现MPS细胞可分泌多达100种以上的酶类和其他生物活性物质。这些物质的分子量不一,从分子量仅32的超氧阴离子至分子量达440000的纤维粘连蛋白;功能也各异,参与从促进细胞生长到导致细胞死亡的全部活动。此外,由于MPS细胞的体内分布广泛、可以移动,以及其分泌产物作用的多样性,这种分泌活性具有重要的生理与病理意义。表9-3列出了MPS细胞所分泌的几类主要产物,MPS细胞的许多功能都有赖于这些分泌产物的参与。
表9-3 单核吞噬细胞系统细胞的分泌产物
种类 |
实例 |
补体 |
C1~C9,B、D、I、H因子,C1抑制物 |
凝血因子 |
V、VⅡ、ⅠX、X因子,凝血酶原 |
酶类 |
各种中性蛋白酶,酸性水解酶,溶菌酶 |
生物活性酯类 |
花生四烯酸衍生物(前列腺素、白三烯等) |
激素样物质 |
VitD3,促肾上腺皮质激素,β内啡肽 |
细胞因子 |
IL-1、IL-3、IL-6,TFN-a,IFN-a,-β、FGF |
反应性中间产物 |
活性氧(1O2、H2O2、OH-,O2-),亚硝酸盐 |
其他 |
多种生长因子及刺激因子,嘌呤及嘧喧产物,各种结合蛋白、连接蛋白及酶抑制物等 |
FGF:纤维细胞生长因子
四、单核吞噬细胞系统细胞的功能
MPS细胞具有重要的生物作用,不仅参与非特异性免疫防御,而且是特异性免疫应答中一类关键的细胞,广泛参与免疫应答、免疫效应与免疫调节。
(一)免疫防御功能
病原微生物侵入机体后,在激发免疫应答以前即可被MPS细胞吞噬并清除,这是机体非特异免疫防御机制的重要环节。由于其吞噬能力较强,故有人将MPS细胞称为机体的清道夫。在致病微生物激发机体产生特异性抗体后,覆盖于病原体表面的IgG及补体激活片段C3b可与MPS细胞表面的FcR及CR1结合,发挥调理作用,使病原体更易被吞噬。被吞入的细菌可被细胞内的某些酶类或活性氧所杀灭;另一方面,在对异物颗粒的吞噬、杀灭过程中,可能出现酶体外漏现象,从而造成对邻近正常组织的损伤。
(二)免疫处自稳功能
机体生长、代谢过程中不断产生衰老与死亡的细胞以及某些衰变的物质,它们均可被单核吞噬细胞吞噬、消化和清除,从而维持内环境稳定。
(三)免疫监视功能
MPS细胞构成机体肿瘤免疫的重要一环。一般认为只有激活的巨噬细胞才能有效地发挥杀瘤效应,其机制可能是:①吞噬肿瘤细胞;②借助抗瘤抗体的ADCC作用杀伤瘤细胞;③产生TNf 及IL-1等细胞因子,直接或间地发挥杀瘤作用;④产生某些酶及活性氧分子直接杀伤或抑制肿瘤细胞生长。
(四)抗原呈递功能
MPS细胞是最重要的一类抗原呈递细胞。外来抗原经单核吞噬细胞处理后呈递给T细胞,这是诱发免疫应答的先决条件。此外,在抗原呈递过程中MPS细胞产生的IL-1也是TH活化不可缺少的刺激信号。
(五)免疫调节功能
MPS细胞在免疫调节中发挥重要的作用。由于激活程度及分泌产物的不同,MPS细胞的免疫调节作用有双相性;另一方面,体内各种因素也可通过影响单核吞噬细胞的膜分子表达等途径调节MPS细胞功能状态。
1.正相调节作用 MPS细胞可通过下列途径启动和增强免疫应答,包括:①抗原呈递作用,诱导免疫应答启动;②分泌具有免疫增强作用的各类生物活性物质,如IL-1、TNF-a、补体成分、各类生长因子等。
2.负相调节作用巨噬细胞过度激活可成为抑制性巨噬细胞,后者可分泌多种可溶性抑制物如前列腺素、活性氧分子等,抑制淋巴细胞增殖反应或直接损伤淋巴细胞。
3.体内各种因素通过调控MPS细胞功能状态而发挥免疫调节作用多种神经肽及激素样物质如P物质、皮质激素、性激素等均可通过相应受体而调控巨噬细胞的功能状态;另外,某些神经肽与细胞因子(如IL-1、TNF-a、IFN-γ)可诱导巨噬细胞MHC-Ⅱ类抗原的表达,从而调控其抗原呈递功能。
(六)其他功能
MPS细胞还广泛参与炎症、止血、组织修复、再生等过程。
第二节 其他抗原呈递细胞
除了MPS细胞外,体内还有其它某些细胞具有原呈递功能。
一、树突状细胞
树突状细胞(dendritic cell,DC)是体内不同于MPS细胞的另一类重要的抗原呈递细胞,其共同的生物学特性是细胞表面有许多树枝状突起,胞内具有丰富的线粒体,但粗面内质网、溶酶体与核糖体不发达,细胞表面无绵羊红细胞受体及SmIg。DC在混合淋巴细胞反应中是重要的刺激细胞。一般认为DC无吞噬功能,但由于其表面表达较高密度的MHC-Ⅱ分子,且具有树突状突起,表面积较巨噬细胞更大,有利于接触抗原并呈递给T细胞,故DC在抗原呈递中发挥重要作用。来源于骨髓前体细胞的DC分布于外周血或各类淋巴组织中,一般只占所在器官全部细胞的1%以下。由于所居留的组织部位不同或处于不同的发育阶段,DC可有不同的名称,并表现出某些特有的生物学特征(表9-4)。
表9-4 各类DC的主要生物学特征
类别 |
体内分布 |
MHC-Ⅱ |
FcR |
C3R |
Birbeck颗粒 |
滤泡树突状细胞 |
淋巴滤泡 |
+/? |
+ |
+ |
- |
淋巴样树突状细胞 |
淋巴组织 |
++++ |
- |
-/+ |
- |
并指状细胞 |
淋巴组织 |
++++ |
- |
- |
- |
朗格罕细胞 |
表皮及某些上皮 |
++++ |
+ |
+ |
+ |
1.滤泡树突状细胞(folliculardendritic cell,.FDC) FDC定居于淋巴结浅皮质区淋巴滤泡生发中心内。FDC与抗原抗体复合物高度亲合力,能够捕获和滞留抗原,并在记忆B细胞发育中起重要作用,是参与再次免疫应答的APC。
2.淋巴样树突状细胞(Lymphoiddendritic cell,LDC) LDC主要分布在淋巴结和脾内,在移植排斥反应中起重要作用。
3.并指状细胞(interdigitatingcell,IDC)IDC主要定位于淋巴组织胸腺依赖区,可能由皮肤Langerhans细胞移行而来。在淋巴组织中,IDC的星状突起插入其它细胞之间,故命名为并指状细胞。它可能是淋巴结中主要的APC,并对抗原特异性T细胞具有很强刺激作用。
4.朗格罕细胞(Langerhanscell,LC) LC位于表皮和胃肠上皮,其特征性胞内结构是胞浆中的柱状Birbeck颗粒,该颗粒参与LC抗原呈递作用的各个环节。LC是定居在皮肤中的APC,约占皮肤细胞总数的5%~10%,在介导接触性皮肤超敏反应中起关键作用。
二、B细胞
正常B细胞及B淋巴瘤细胞系均表达MHC-Ⅱ类抗原,也属于抗原呈递细胞。与B细胞抗原呈递功能相关的膜分子是SmIg,后者与抗原具有高亲和力,从而可以高度特异性地浓缩非已抗原,并以与MHC-Ⅱ类分子结合成复合体的形式将抗原呈递给TH。故尤其对低度的抗原,B细胞是一类有效的APC。
三、内皮细胞及其他抗原呈递细胞
人的内皮细胞也表达MHC-Ⅱ类抗原并可与TH细胞相互作用,故也是一种APC,它在介导迟缓型超敏反应中发挥重要作用。此外,某些上皮细胞、神经胶质细胞、间质细胞等在静止状态下或在γ干扰素的诱导下可表达MHC-Ⅱ类抗原,并具有APC的功能。
病毒感染的细胞以及肿细胞等作为细胞免疫效应的靶细胞可将非已抗原呈递给细胞毒性T细胞,因此从广义上讲这些靶细胞也属于抗原呈递细胞。
第三节 抗原呈递细胞的抗原呈递作用
细胞在其表面以能被T细胞受体(TcR)特异性识别的方式表达抗原的过程称为抗原呈递。APC以MHC-Ⅱ分子限制的方式将抗原递给辅助性T细胞(TH);各种靶细胞心以MHC-Ⅰ限制的方式将抗原呈递给细胞毒性T细胞(TC)。APC的抗原呈递作用是一个涉及抗原摄取、处理与呈递的复杂过程。各类APC呈递抗原的机制不尽相同,本章主要以巨噬细胞对非已抗原的呈递为例叙述。
一、抗原摄取
非已抗原(如细菌抗原或可溶性抗原)以非特异性方式与巨噬细胞结合,后者借助吞噬、胞饮、吸附或FcR、CR1介导的调理作用摄取抗原。这个过程也称为内吞作用(endocytosis)。
二、抗原加工
T细胞通常不能识别天然蛋白抗原的决定簇。被APC摄入的天然抗原须在胞内通过代谢而修饰成能与MHC-Ⅱ类分子结合的、具有强免疫原性的肽段,此过程称为抗原的处理(antigen processing)。被巨噬细胞摄入的抗原在胞内形成吞噬小体(phagosome),然后与溶酶体融合,形成吞噬酶体(phagolysosome)。后者是一种酸性的囊泡,天然抗原在其内受多种酸性水解酶作用,90%以上的成分被完全裂解为氨基酸并失去免疫原性,10%左右降解为具有α-螺旋结构的免疫原性肽段。后者暴露出与MHC-Ⅱ类分子具有特异亲和力的疏水性残基,并以抗原识别位(agretope)通过非共价键与MHC-Ⅱ类分子肽结合区结合成稳定的螺旋结构。由细胞内粗面内质网合成的MHC-Ⅱ类分子通过与免疫原性多肽的结合,使后者得以避免被完全水解。所形成的多肽MHC-Ⅱ分子复合物可能在高尔基复合体参与下被转运到细胞膜表面TcR识别。
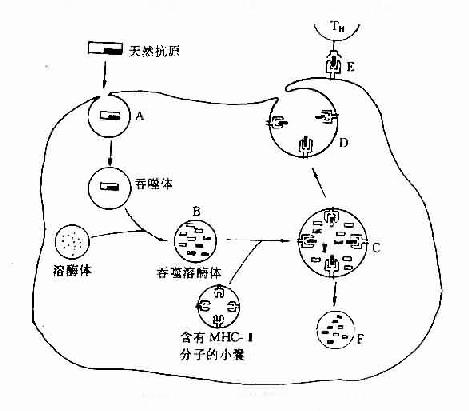
图9-1 巨噬细胞对抗原的摄取、加工及呈递
A.内吞
B.天然抗原在吞噬溶酶体中降解;
C.免疫性多肽与MHC-Ⅱ分子结合复合物;
D.多肽-MHC-Ⅱ分子复合物的转动与表达;
E.TH的TCR联合识别多肽-MHC-Ⅱ分子复合物;
F.未与-MHC-Ⅱ分子结合的肽链彻底降解
三、抗原呈递
表达于巨噬细胞表面的多肽-MHC-Ⅱ分子复合物被呈递给TH细胞。供TcR识别的先决条件是两种细胞的直接接触并相互作用。这种细胞间的相互作用涉及APC与TH表面多种分子。除了TcR特异性地同时识别多肽-MHC-Ⅱ分子的复合物外,某些粘附分子也参与抗原呈递过程。例如TH表面的淋巴细胞功能相关抗原1(lymphocytefunctiom-associatedantigen-1,LFA-1)、CD2和CD4分子可分别与APC表面的细胞间粘附分子1(ICAM-1)、淋巴细胞功能相关抗原3(LFA-3)和MHC分子形成共轭物,并促进TcR与多肽-MHC分子复合物的相互作用。此外,在TcRαβ链识别抗原过程中,TH细胞表面的CD3分子参与刺激信号的传递并介导T细胞产生淋巴因子及发挥效应功能。
由上可见,APC呈递抗原供TcR识别并导致T细胞激活是一个受到严格调节的复杂过程。其中某些细节尚未弄清,有待进一步探讨。
